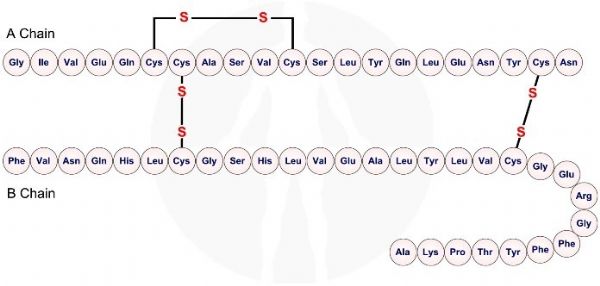
האינסולין בנוי משתי שרשרות פפטידיות, אחת קרויה שרשרת A (chain A) אשר בנויה מ - 21 חומצות אמינו, והשניה קרויה שרשרת B (chain B) ובנויה מ - 30 חומצות אמינו. שתי השרשרות מקושרות על ידי שני גשרים ביסולפידיים. גשר ביסולפידי שלישי מצוי על שרשרת A בלבד.
אינסולין הינו הורמון פפטידי ( המורכב מחומצות אמינו ) המיוצר באזור בלבלב הנקרא איי לנגרהנס. תפקידו הראשי של אינסולין הוא לווסת את רמת הגלוקוז בדם. תאי הלבלב רגישים לעליית הגלוקוז בדם ומפרישים אינסולין אל הדם. אינסולין משפיע על תאי השרירים ורקמת השומן וגורם לתאים אלו לייצר ולהפריש יותר נשאי גלוקוז אשר מכניסים גלוקוז אל התאים מהדם. אינסולין גורם לכניסת חומרים נוספים אל התאים, כגון: יוני אשלגן, קריאטין וחומצות אמינו.
מלבד תפקידו המטבולי, לאינסולין יש תפקיד אנבולי אשר חשוב לגדילה באופן כללי. אינסולין משפיע על קצב חלוקה והתמיינות תאים.
ייצור אינסולין
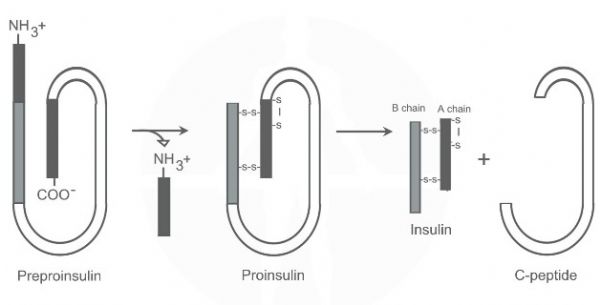
המידע האצור ב – DNA אינו מקודד ליצירת אינסולין באופן ישיר, אלא לייצור חלבון בשם פרה-פרואינסולין - preproinsulin. הפרה-פרואינסולין עובר עיבוד בתאי הבטא לפרואינסולין – proinsulin. מולקולת הפרואינסולין מתפרקת לשני חלבונים: אינסולין ולפפטיד - C-peptide. אינסולין ופפטיד C מאוכסנים בגרנולות הפרשה אשר בעת הצורך ינועו אל עבר ממבראנת התא יתמזגו עמה והאינסולין יופרש.
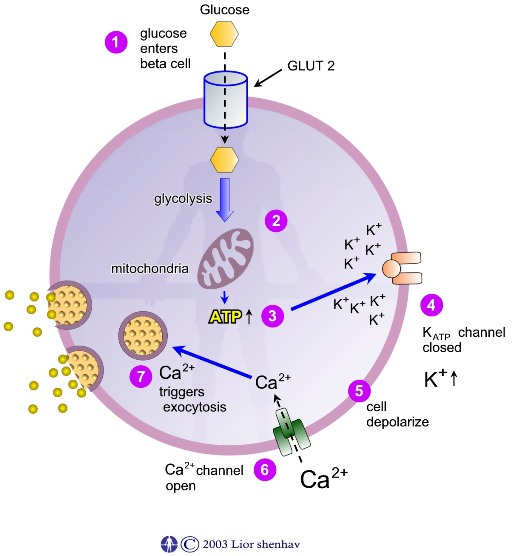
4. מולקולות ATP נקשרות לתעלות אשלגן רגישות ל – ATP וגורמות לסגירתן. סגירת תעלות האשלגן גורמת לעליה ברמת יוני האשלגן +K בתא.
5. עליה ברמת יוני האשלגן +K בתא גורמת לשינוי המטען החשמלי - דפולריזציה בתאי הבטא.
6. דפולריזציית התא גורמת לפתיחת תעלות סידן ולעליה ברמת יוני הסידן +Ca2 בתא.
7. עליה ברמת יוני הסידן +Ca2 גורמת לגרנולות ההפרשה אשר מאכסנות אינסולין לנוע אל עבר ממבראנת התא, להתמזג עם ממבראנת התא ולהפריש את האינסולין אל מחוץ לתאי הבטא.
אינסולין מופרש בכמות גדולה יותר כאשר גלוקוז ניתן באופן פומי לעומת מתן ורידי
כאשר גלוקוז ניתן באופן פומי – דרך מערכת העיכול, מופרשים בתגובה לאכילה הורמוני מעי בשם אינקרטינים - בעיקר GLP1 - Glucagon-Like Peptide 1. הורמונים אלו גורמים לעליה ברמת ה – cAMP – מעביר משני בתא, אשר משפיע באופן סינרגיסטי עם יוני הסידן על גרנולות ההפרשה אשר מכילות אינסולין.
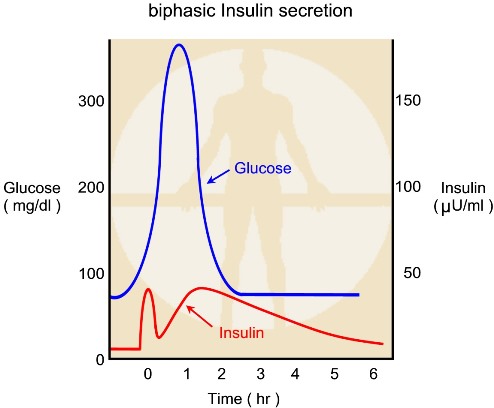
הפרשת אינסולין הינה בי פאזית
אינסולין אינו מופרש כאשר רמת הסוכר בדם הינה מתחת ל – 50 מ"ג / דצ"ל.
מחצית מכמות האינסולין המקסימלית מופרשת כאשר רמת הגלוקוז הינה כ – 150 מ"ג / דצ"ל. כמות האינסולין המקסימלית מופרשת כאשר רמת הגלוקוז בדם הינה מעל 300 מ"ג / דצ"ל.
בתגובה לגלוקוז אינסולין מופרש בשתי פאזות. בפאזה הראשונה מופרש אינסולין בין 5 – 15 דקות, לאחר מכן מתחילה עליה הדרגתית ברמת האינסולין המופרשת, כל זמן שרמת הגלוקוז בדם גבוהה.
הפרשת האינסולין בשתי הפאזות אינה רציפה. אינסולין מופרש בכמות מסוימת עם תנודות - עליות וירידות כל 3 – 6 דקות.
אינסולין מופרש אל הדם כהורמון חופשי
הלבלב מפריש אינסולין כמות השווה ל- 1 מ"ג ליום, או 40 מק"ג / 1 יחב"ל - IU לשעה.ריכוז האינסולין בדם הפורטלי של 2 עד 4 נ"ג / מ"ל – ng/mL או 50 – 100 מק"ל / מ"ל.
יותר ממחצית מכמות האינסולין מפונה על ידי הכבד, כך שריכוז האינסולין הינו כ- 0.5 נ"ג / מ"ל – ng/mL או כ- 12 מק"ל / מ"ל.
זמן מחצית החיים של אינסולין המוזרק לווריד בדם הינו בין 5 עד 6 דקות. זמן מחצית החיים עשוי להתארך במקרה של חולי סוכרת בהם קיימים נוגדנים לאינסולין.
אינסולין מפורק בשלוש רקמות עיקריות: כבד, כליות ושרירים. לפחות 50% מהאינסולין שמגיע לכבד דרך הווריד הפורטל נלקח במעבר אחד ואינו מגיע למחזור הדם ההיקפי. בכליות אינסולין מסונן על ידי נפרוני הכליה וחלקו נהרס בתהליך. ירידה חדה בתפקוד הכליות משפיעה על קצב פינוי האינסולין בדם.
קולטן האינסולין - Insulin receptor
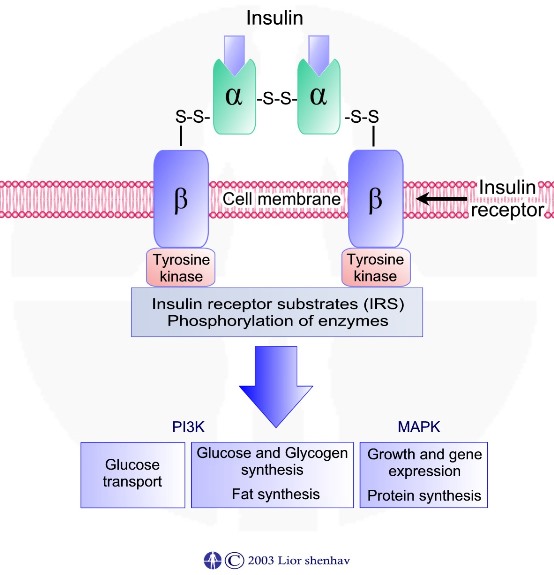
קולטן האינסולין בנוי משתי יחידות אלפא אשר נמצאות מחוץ לתא ושתי יחידות בטא אשר חוצות את ממבראנת התא. שתי היחידות קשורות בניהן בקשרים די סולפידיים. קשירת אינסולין ליחידות האלפא גורמת לשינוי מבני אשר בסופו של דבר גורם לזרחון שיירי טירוזין אשר קשורות ליחידות הבטא.
זרחון שיירי הטירוזין גורם להתחלת שרשרת תהליכים אשר כוללת: זרחון חלבונים נוספים בתא החל מקבוצת חלבונים – IRS - insulin-receptor substrates והמשך זרחון חלבונים במסלולים שונים בתא.
בנוסף להשפעתו על סינתזת שומנים, אינסולין משפעל את האנזים ליפופרוטאין ליפאז - Lipoprotein lipase, אשר גורם לפירוק טריגליצרידיםמכילומיקרונים ו - VLDL ואחסונם בתאי שומן.
אינסולין מעכב ליפוליזה על ידי עיכוב ליפאז – HSL ומגביר כניסת יוני אשלגן +K לתא.
בנוסף להשפעת אינסולין על כניסת גלוקוז אל תאי השריר, אינסולין מעודד כניסת חומצות אמינו לתא. הגברת סינתזת חלבונים והפחתת פירוק חלבונים. אינסולין מגביר את חדירות תאי השריר לגופי קטון, יוני אשלגן לתא +K, קריאטין וקארניטין.
כבד
אינסולין מעודד כניסת גלוקוז אל תאי הכבד וזרחון מולקולות גלוקוז על ידי גלוקוקינאז. אינסולין משפעל את האנזים גליקוגן סינתאז ומולקולות הגלוקוז מופנות למסלול סינתזת גליקוגן.
אינסולין מעכב את תהליך הגלוקונאוגנזה, מאחר אין צורך ליצור גלוקוז. אינסולין מעכב יצירת גופי קטון, מאחר ואינסולין מעודד סינתזת שומנים ומעכב את פירוקם.
אינסולין מעודד סינתזת חלבוניםבכבד.
כליות
אינסולין משפיע על הסינון הכלייתי של יוני נתרן Na+. אינסולין גורם לכליות לספיגה חוזרת של יוני נתרן, שימור נתרן בגוף ואגירת נוזלים.
הלבלב האקסוקריני
בתאים האצינריים בלבלב – אינסולין מגביר ייצור אנזימי עיכול: אלפא עמילז, טריפסין וכימוטריפסין. מקור עיקר ההגברה ביצירת האנזימים הוא בהגברת התגובה לאצטיל-כולין ולכוליציסטוקינין – CCK.
קיבה
אינסולין גורם לשחרור יוני מימן בקיבה.
גונדות
אינסולין גורם להתרבות קולטנים ל FSHולLH- על תאי הגרנולוזה בשחלה ועל תאי ליידיג באשך. בחוסר אינסולין מופחתת סינתזת הורמוני המין בתאים אלו.
תאי אנדותל
בתאי אנדותל אינסולין מאיץ קליטת חומצה אסקורבית – ויטמין C. מחסור בחומצה אסקורבית בתאי האנדותל מאיץ הפרוק ומדכא יצור של גלוקוזאמינוגליקן - Glucoseaminoglycan המהווה את המטריקס המצמיד את תאי האנדותל לדופן כלי הדם. מחסור באינסולין גורם לנשירת תאי אנדותל מדופן כלי הדם ומתהווים מוקדים טרשתיים בכלי הדם.
השפעת אינסולין בתא
1. מעבר נשאי גלוקוז אל ממברנת התא לשם הכנסת גלוקוז אל התאים.
2. אגירת גלוקוזכגליקוגן.
3. כניסת חומצות אמינו, קריאטין ויונים אל התא.
4. סינתזת חלבונים.
5. סינתזת חומצות שומן.
6. סינתזת DNA וגדילת תאים.
7. עיכוב גלוקונאוגנזה.
8. עיכוב ליפוליזה – פירוק שומן.
9. מעכב אפופטוזיס – מוות תאים.
בקרת פעילות האינסולין תלויה במספר הקולטנים ועוצמת הקשירה אליהם:
רמת האינסולין X עוצמת הקשירה לקולטנים X כמות הקולטנים.
גורמים אשר משפיעים על כמות הקולטנים לאינסולין:
• עליה ברמת הגלוקוז גורמת לעליה בכמות הקולטנים לאינסולין על גבי ממברנות תאי הכבד ורקמת השומן.
• תזונה עתירה בשומן גורמת לירידה בכמות הקולטנים לאינסולין על גבי ממברנות תאי הכבד ורקמת השומן.
• עליה במסת השומן - במצב של השמנה תאי השומן מפרישים כמות גדולה של ההורמון רזיסטין – resistin לפטין, ו-TNF-α – אלו מגבירים את התנגודת לאינסולין.
• עודף באינסולין יגרום להפחתת כמות הקולטנים לאינסולין - down-regulation.
• להורמון פרוגסטרון, אשר מופרש בכמות גדולה בזמן הריון יש השפעה אשר באה לידי ביטוי בהפחתת כמות הקולטנים לאינסולין.
• גיל – תהליך ההזדקנות מלווה בירידה בכמות הקולטנים לאינסולין.
• גלוקוקורטיקואידים – קורטיזון ונגזרותיו, בין שמדובר בעליה טבעית כתוצאה ממצבי דחק או ממתן חיצוני, יגרמו לירידה בכמות הקולטנים לאינסולין.
• תגובה אוטואימונית אשר בה נוצרים נוגדנים כנגד הקולטנים לאינסולין תגרום להרס הקולטנים וירידה במספרם.
סינתוז גליקוגן
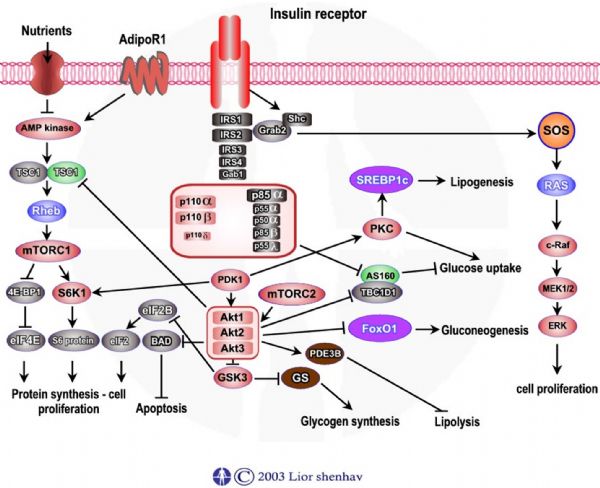
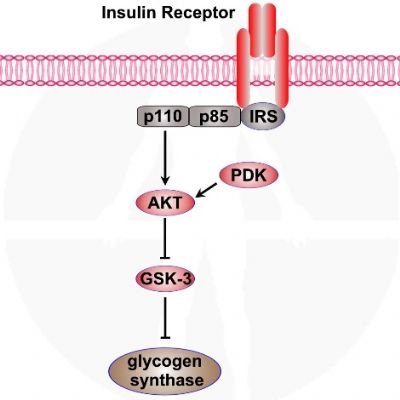
הפעלת הקולטן לאינוסלין גורמת לזרחון גליקוגן סינתאז קינאז – 3 GSK - . זרחון GSK- 3 גורם להפסקת פעילותו של החלבון. כאשר GSK- 3 פעיל גליקוגן סינתאז – Glycogen synthase נמצא במצבו הבלתי פעיל. כך שזרחון GSK- 3 מוביל להפעלת גליקוגן סינתאז – Glycogen synthase ולחידוש מאגרי גליקוגן.
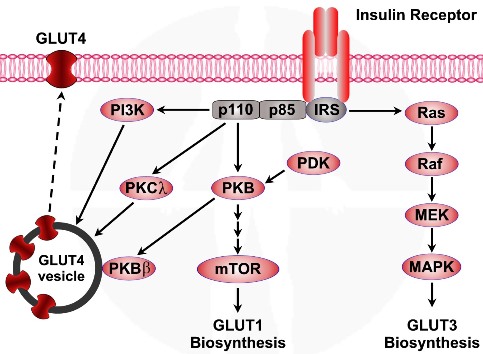
נשא גלוקוז – 4 GLUT נשא גלוקוז – 4 הינו נשא גלוקוז אשר מבוקר עלי יד אינסולין. נשאי גלוקוז – 4 מאוכסנים בתא בתוך ממבראנה פנימית המשמשת ככלי מעבר. עם הפעלת הקולטן לאינסולין מופעלים - PKC, PKB-beta אשר גורמים לדיפוזיה של נשאי הגלוקוז – 4 אל ממבראנת התא.
לפרואינסולין ישנה יעילות ביולוגית בשיעור של כ - 6-15% לעומת האינסולין בהעלאת החדרת גלוקוז לתאים, תלוי בסוג הרקמה. לפרואינסולין אותה יעילות כשל האינסולין במניעת ליפוליזיס. ההשפעה האנטיליפוליטית היא כנראה תוך תאית ולא ממברנאלית ויתכן שמולקולת הפרואינסולין נכנסת לתא המטרה ומומרת בו לאינסולין ע"י אנזימים תוך תאיים.
ככל שעולה חלקו של הפרואינסולין בכלל ההפרשות של תאי הבטא, היעילות ההיפוגליקמית של תאי בטא יורדת.