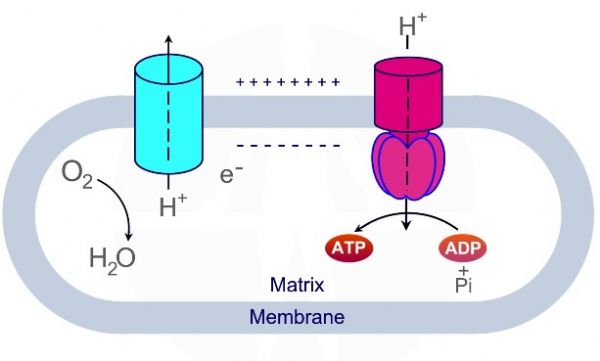
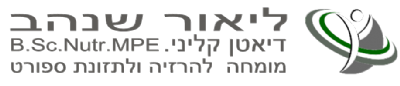העברת האלקטרונים מתבצעת על גבי קומפלקסים 1-4, כאשר קומפלקסים 1,3,4 משמשים כמשאבות פרוטונים – מוציאים את הפרוטונים לחלל הבין-ממברנלי. החמצון מתרחש בהדרגה על מנת "לסחוט" כמה שיותר אנרגיה מן האלקטרונים.
ניתן לפשט את התהליך באופן הבא:
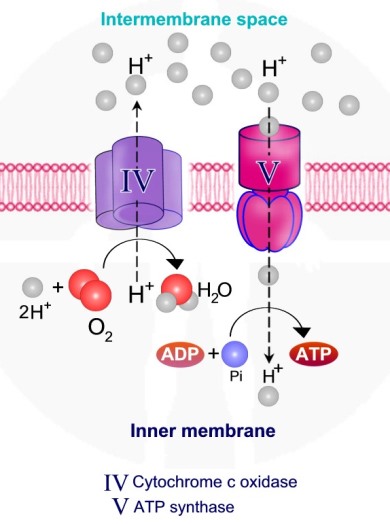
אנרגיית האלקטרונים משמשת לפעולת משאבות פרוטון ולהוצאת פרוטונים אל המרווח הבין ממברנלי., כאשר הפרוטונים נעים במסלול ההפוך דרך אנזים - ATP synthase , האנרגיה האצורה בתהליך משמשת ליצירת ATP.
זרחון חמצוני – שלב ראשון
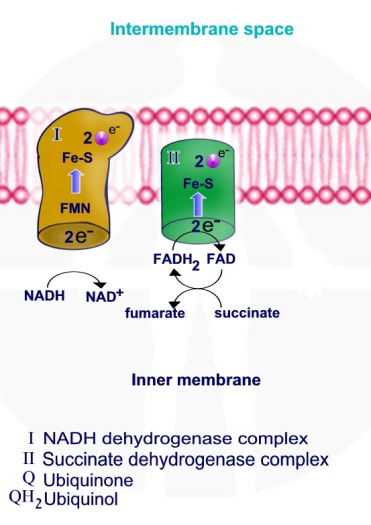
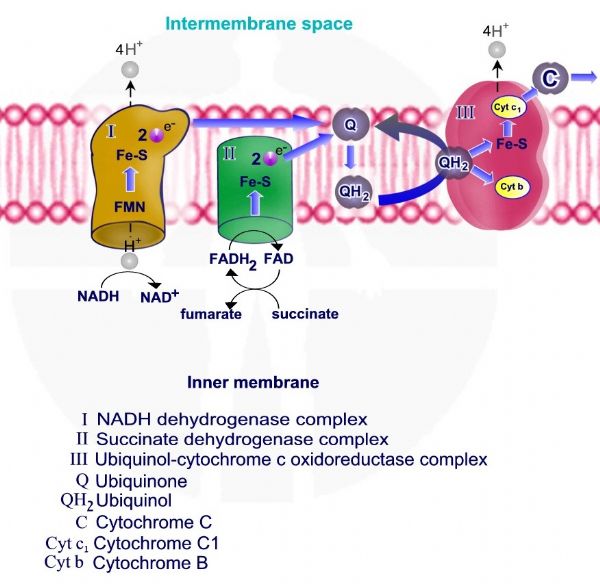
תהליך החמצון הזרחוני מתחיל בחמצון 3 מולקולות NADH ומולקולת FADH2 אשר נוצרות במעגל קרבס. חמצון מולקולות NADH ו – FADH2 גורם להעברת אלקטרונים ממולקולות אלו אל קומפלקס - NADH Hydrogenase - I ואל קומפלקס - Succinate Dehydrogenase - II.
זרחון חמצוני - שלב שני
חמצון מולקולת NADH גורם להוצאת פרוטונים – יוני מימן בעלי מטען חיובי +H דרך קומפלקס I אשר משמש כמשאבת פרוטונים.
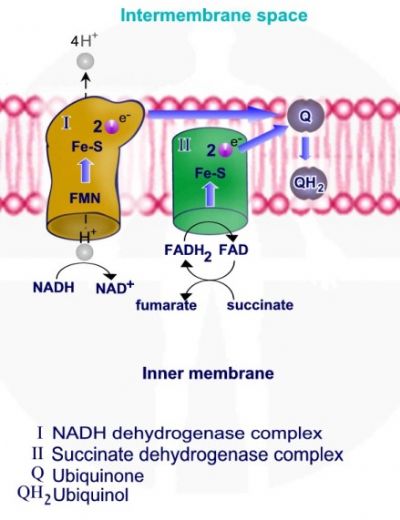
מולקולת NADH מוסרות פרוטון ואלקטרונים דרך קומפלקס I אשר משמש כמשאבת פרוטונים, האלקטרונים מועברים מקומפלקס I ל – Q - קו אנזים קיו 10 - ubiquinone. חמצון מולקולות NADH ל – NAD+ מספק את האנרגיה להוצאת פרוטון אל המרווח הבין ממברנלי.
מולקולות FADH2 מוסרות פרוטונים ואלקטרונים דרך קומפלקס II אשר משמש כמעביר אלקטרונים אך אינו משמש כמשאבת פרוטונים. האלקטרונים מועברים מקומפלקס I ל – Q - קו אנזים קיו 10.
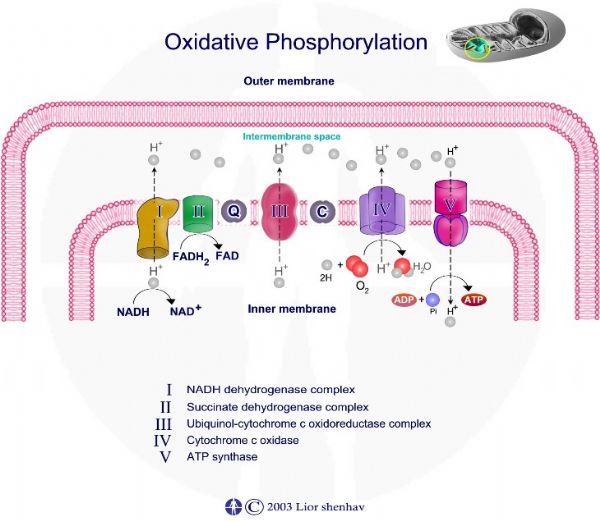
לאחר קבלת אלקטרונים עובר Q - קו אנזים קיו 10 תהליך חיזור ונוצרת מולקולת QH2 – ubiquinol.
QH2 נושא אלקטרונים אך גם פרוטונים ומעביר אותם לקומפלקס III, האלקטרונים אשר מגיעים לקומפלקס III מועברים לציטוכרום C, מעביר האלקטרונים האחרון ומשמשים את קומפלקס III המשמש כמשאבת פרוטונים להוצאת הפרוטונים אל המרווח הבין ממברנלי.
חיזור ubiquinone - Q ל – ubiquinol – QH2 דורש שני אלקטרונים ושני פרוטונים ומתרחש בשני שלבים.
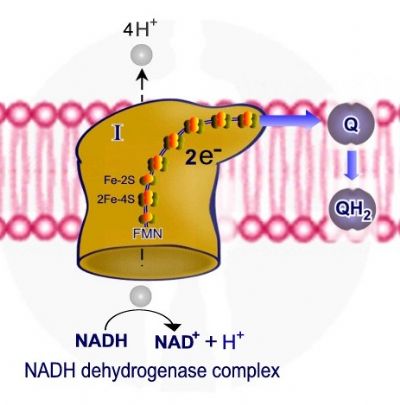
קומפלקס - NADH Hydrogenase /NADH-coenzyme Q oxidoreductase I , מאפשר מעבר שני אלקטרונים ממולקולת NADH ל – ubiquinone – Q. האלקטרונים מועברים דרך מולקולת flavin mononucleotide -FMN ודרך שישה מרכזי ברזל – נחושת, שני מרכזי Fe-2s וארבעה מרכזי 2Fe – 4s.
מעבר האלקטרונים מאפשר מעבר ארבעה פרוטונים אל המרווח הבין ממברנלי. בסוף התהליך האלקטרונים מועברים ל - ubiquinone – Q אשר עובר חיזור והמרה ל QH2 – ubiquinol.
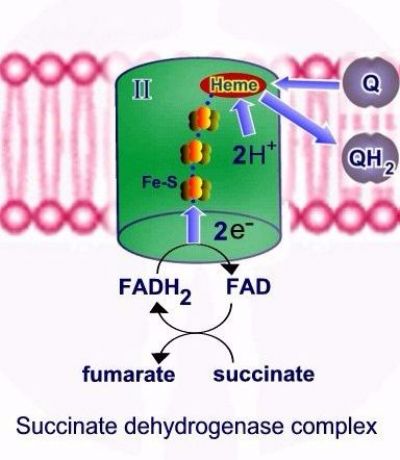
קומפלקס II אינו משמש כמשאבת פרוטונים, האלקטרונים והפרוטונים מועברים ל Q10 ומשם מועברים לקומפלקס Q-cytochrome c oxidoreductase -III.
זרחון חמצוני - שלב שלישי
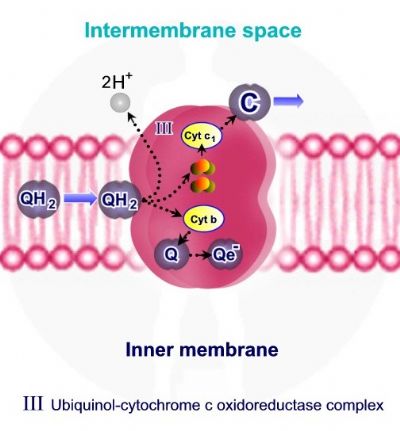
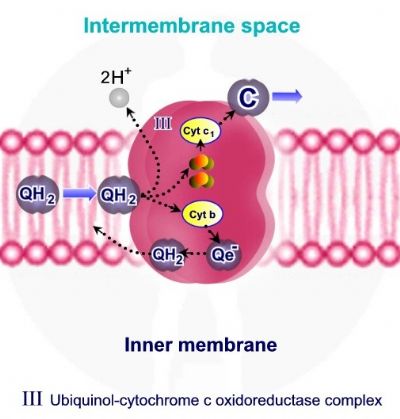
קומפלקס Q-cytochrome c oxidoreductase – III מעביר אלקטרונים מ QH2 אשר משמש כנשא אלקטרונים ופרוטונים מקומפלקס I ומקומפלקס II אל ציטוכרום C, תוך כדי הוצאת 4 פרוטונים אל המרווח הבין ממברנלי.
קומפלקס Q-cytochrome c oxidoreductase – III מכיל 2 קבוצות Fe-2S, ושני ציטוכרומים – חלבונים בעלי קבוצות הם - Heme. ציטוכרום – Cyt C1 אשר מכיל קבוצת Heme אחת וציטוכרום Cyt B, אשר מכיל שתי קבוצות Heme.
מחזור Q מחזור Q מתאר את פריקת האלקטרונים והפרוטונים מ - QH2 על ידי קומפלקס III. המחזור בעל שני שלבים:
שלב ראשון • QH2 אשר מגיע מקומפלקס I ומקומפלקס II, נקשר אל קומפלקס III. • שני פרוטונים משתחררים מ - QH2 ונפלטים אל המרווח הבין ממברנלי. • שני אלקטרונים משתחררים מ - QH2, אלקטרון אחד עובר במסלול Fe-2S ,אל ציטוכרום – Cyt C1 ואל ציטוכרום C. היות ציטוכרום C מסוגל לקשור אלקטרון אחד, האלקטרון השני אשר משתחרר מ - QH2 צריך לעבור מסלול שונה. • האלקטרון השני עובר דרך ציטוכרום Cyt B ומשם בחזרה ל – Q. • קשירת אלקטרון אחד ל – Q יוצרת מצב ביניים בו נוצר רדיקל -Semiqiunone – Qe.
שלב שני • בשלב השני של מחזור ה – Q מגיעה מולקולה חדשה של - QH2 ונקשרת לקומפלקס III. • כמו בשלב הראשון של המחזור, שני פרוטונים נפלטים אל המרווח הבין ממברנלי ושני אלקטרונים משתחררים מ - QH2. • אלקטרון ראשון עובר במסלול Fe-2S ,אל ציטוכרום – Cyt C1 ואל ציטוכרום C. • האלקטרון השני עובר דרך ציטוכרום Cyt B, אך להבדיל מהשלב הראשון, אלקטרון זה עובר למולקולת Semiqiunone – Qe- ( אשר נצורה בשלב הראשון ) ומחזר אותה ל - QH2. מולקולת ה - QH2 יוצאת מהקומפלקס וחוזרת אליו.
משמעות המחזור – רק אלקטרון אחד מבין שני האלקטרונים אשר משתחררים מ - QH2, יכול לעבור במסלול במסלול Fe-2S ,אל ציטוכרום – Cyt C1 ואל ציטוכרום C. מחזור ה – Q מאפשר את "שמירת" האלקטרון השני ומחזורו, כך שמסלול הובלת האלקטרונים אינו מאבד אלקטרון.
זרחון חמצוני – שלב רביעי
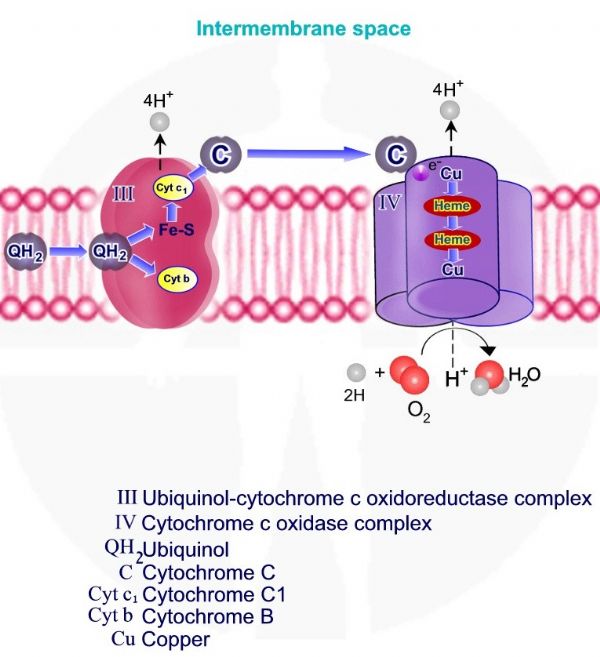
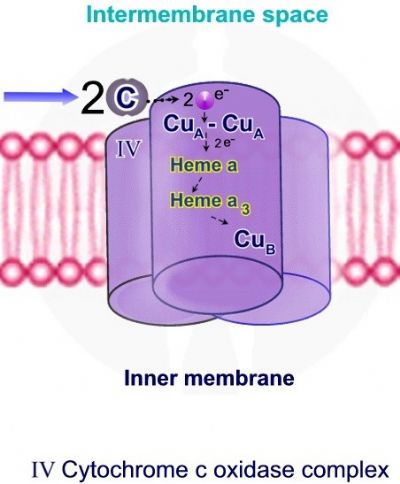
ציטוכרום C, שעבר חיזור על ידי קומפלקס III מעביר את האלקטרון לקומפלקס האחרון בשרשרת הובלת האלקטרונים – קומפלקס Cytochrome c oxidase – IV. בתהליך נפלטים ארבעה פרוטונים אל המרווח הבין ממברנלי והאלקטרונים משמשים לקשירת חמצן מולקולרי למימן ליצירת מולקולות מים.
קומפלקס Cytochrome c oxidase – IV מכיל 2 קבוצות חשובות - 2 קבוצות הם Heme ואטומי נחושת. קבוצות הם Hema: קומפלקס 4 מכיל שתי קבוצות הם: Heme A ו – Heme A3. אטומי נחושת Copper - Cu: קומפלקס IV מכיל 3 אטומי נחושת. שני אטומי נחושת - CuA יוצרים מרכז – CuA-CuA ואילו אטום נחושת שלישי - Cub יוצר מרכז נוסף עם Heme A3, אשר משמש לחיזור חמצן מולקולרי וליצירת מולקולת מים.
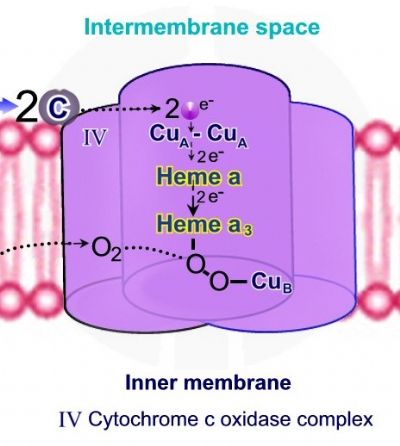
התהליך מתבצע במספר שלבים. שלב 1 - 2 מולקולות של ציטוכרום C נקשרות אל קומפלקס IV, כל מולקולת ציטוכרום C משחררת אלקטרון אחד. האלקטרונים מועברים אל מרכז CuA-CuA, Heme A – Heme A3 ואל אטום הנחושת האחרון Cub. קליטת אלקטרונים גורמת לחיזור Heme A3 ויון הנחושת ממצב Cu+2 ל – +Cu. כאשר Heme A3 ו – Cub נמצאים במצבם המחוזר, קיימת היכולת לקשירת חמצן מולקולרי - O2.
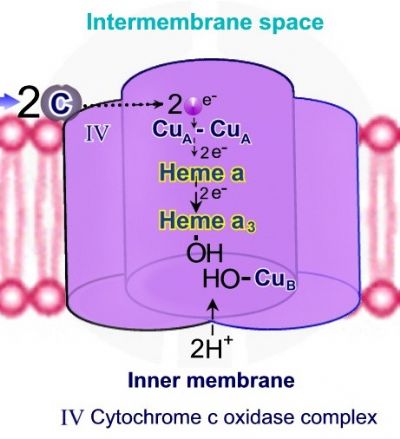
שלב 2 – קשירת חמצן מולקולרי יוצרת גשר פרוקסידי בין Heme A3 ל – Cub . שתי מולקולות חדשות של ציטוכרום C נקשרות אל קומפלקס IV, ושני אלקטרונים נוספים עוברים בין הקבוצות השונות עד לקשירת אלקטרון אחד ל - Heme A3ואלקטרון שני ל – Cub. שני פרוטונים מתוך המיטוכונדריה נכנסים אל קומפלקס IV ומשמשים לביקוע הקשר בין אטומיהחמצן לקבלת שתי קבוצות הידרוקסיד - -OH, האחת קשורה ל - Heme A3 והשניה קשורה ל – Cub.
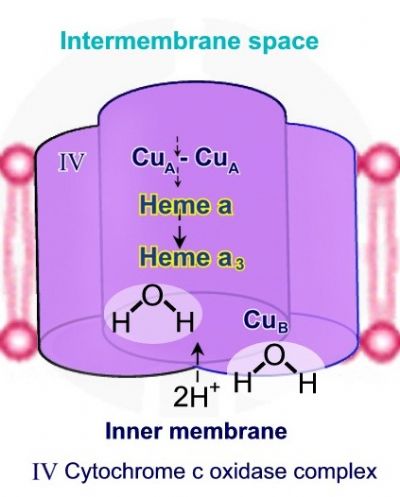
שלב 3 – שני פרוטונים נוספים מתוך המיטוכונדריה נכנסים אל קומפלקס IV, נקשרים אל קבוצות ההידרוקסיד – -OH ליצירת שתי מולקולות מים H-O-H.
זרחון חמצוני שלב חמישי
השלב החמישי בתהליך הזרחון החמצוני הינו שלב יצירת ATP. לאורך תהליך העברת האלקטרונים, פרוטונים נפלטו אל המרווח הבין ממברנלי על קומפלקס I, III ו – IV. ריכוז גבוה של פרוטונים בעלי מטען חיובי +H, גורם לשינוי המטען ולירידה ב - pH - גרדיאנט חשמלי וגרדיאנט כימי - בין האזור הבין ממברנלי - חיובי ולבין תוך המיטוכונדיה – המטריקס.
שני הגרדיאנטים שנוצרים יוצרים כוח אשר מושך את הפרוטונים אל המטריקס – Proton motive force. פרוטונים יכולים לחזור אל המטריקס דרך האנזים - ATP synthase הידוע גם כקומפלקס V.
תנועת הפרוטונים דרך ATP synthase, גורמת לרוטציה באנזים ולקשירת קבוצת פוספט – Pi ל –ADP וליצירת ATP.
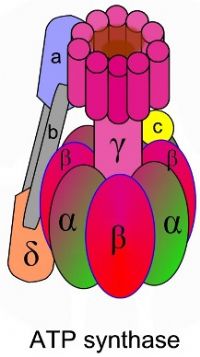
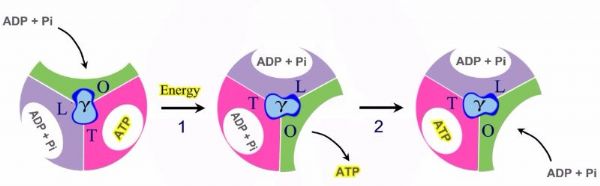
קומפלקס V – ה- ATP Synthase קומפלקס V בנוי כמנוע לייצור ATP. הקומפלקס בנוי מיחידות אשר חוצות את הממברנה ותפקידן להעביר פרוטונים אל תוך המיטוכונדריה מהחלל הבין ממברנלי. הקומפלקס מכיל חלק מסתובב, אשר בנוי מ-6 תתי יחידות, 3 יחידות אלפא ו – 3 יחדות בטא. כאשר פרוטונים מועברים דרך הקומפלקס, עבור כל סיבוב של הקומפלקס נוצרות 3 יחידות ATP.
תת יחידה – גמא, מאחדת בין יחידות האלפא והבטא, לקבלת אתר פעיל לייצור ATP. כך מתקבלים שלושה אתרים פעילים באנזים בעלי 3 קונפורמציות ברוטציה: Loose, Tight, Open
אל ה- Loose נכנס ADP+Pi., בעזרת הסיבוב ואנרגיה הופך ל- Tight, מאפשר את יצירת ה – ATP, מעבר הפרוטונים הופך ל- Open ומשחרר את ה- ATP.
מעכבים
מפרי צימוד:
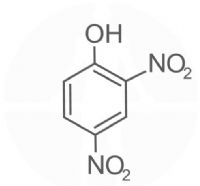
די ניטרופנול - 2,4-Dinitrophenol.
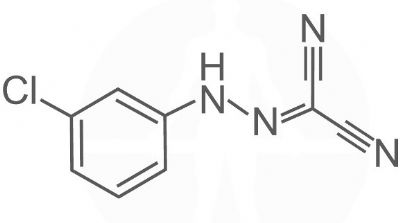
Carbonyl cyanide m-chlorophenyl hydrazine – CCCP.
מפרי צימוד גורמים להפרעה במפל האלקטרוכימי על ידי הכנסת יוני +H בחזרה אל המרווח התוך ממברנלי. מאחר וסינתזת ATP תלויה במעבר של יוני +H בחזרה אל המרווח בתוך ממברנלי דרך ATP Synthase, מעבר יוני +H בחזרה אל תוך המיטוכונדריה בדרך עקיפה ימנע את סינתזת ה – ATP.
די ניטרופנול -
2,4-Dinitrophenol.
Carbonyl cyanide m-chlorophenyl hydrazine – CCCP
מעכב ATP Synthase:
אוליגומיצין נקשר אל היחידה F0 ומונע מעבר יוני H דרך ATP Synthase.
Oligomycin A
קולטי אלקטרונים:
קולטי אלקטרונים קוטעים את רצף מעבר האלקטרונים בשרשרת העברת האלקטרונים אשר מייצרת את האנרגיה הדרושה לשאיבת יוני ה – H אל המרווח הבין ממברנלי. הפרעה ביציאת יוני ה – H אל המרווח הבין ממברנלי תגרום להפרעה ביצירת הגרדיאנט האלקטרוכימי ותגרום להפסקת ייצור ATP.
רוטנון – Rotenone, קולט אלקטרונים המועברים מקומפלקס I אל ubiquinone – Q, על ידי קשירה לאתר הפעיל של ubiquinone – Q.
Rotenone
Antimycin A
אנטימיצין – Antimycin A, נקשר אל קומפלקס Q-cytochrome c oxidoreductase – III ומונע חמצון ubiquinol.
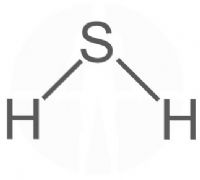
מימן גופריתי – Hydrogen sulfide – HS.
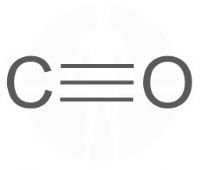
פחמן חד חמצני – Carbon monoxide – CO.
אזיד – Azide.
ציאניד – Cyanide.
הינם קולטי אלקטרונים אשר מונעים את קשירת החמצן למרכזי Fe – Cu
בקומפלקס Cytochrome c oxidase – IV, מונעים את חיזור החמצן.
מעבורות
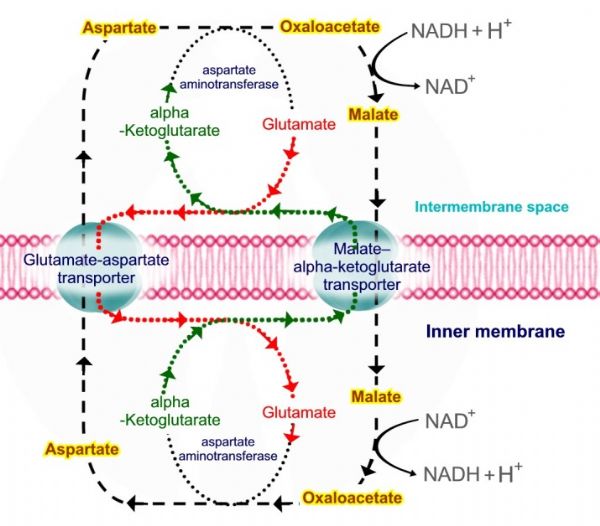
מעבורת מלאט – אספרטט - malate-aspartate shuttle.
פעילה בעיקר בכבד ובלב, מעבירה אלקטרונים ללא הפסד אנרגיה.
קומפלקס - NADH Hydrogenase – I, יכול לקבל אלקטרונים אך ורק ממולקולות NADH אשר נמצאות במטריקס של המיטוכונדריה. מאחר וקיים צורך בחמצון מולקולות NADH ל – +NAD בציטוזול, קיים הצורך בהעברת האלקטרונים לשרשרת הובלת האלקטרונים בדרך עקיפה.
אוקסלואצטט בציטופלזמה מומר למלאט - Malate על ידי האנזים מלאט דהידרוגנאז – malate dehydrogenase, התהליך יוצר מולקולה טעונה באלקטרונים אשר מתקבל מהמרת +NADH + H ל – +NAD, מלאט נכנס אל המיטוכונדריה על ידי טרנספורטר – Malate – alpha ketoglutarate transporter, אשר מוציא אלפא קטוגלוטראט בכיוון הנגדי.
בתוך המיטוכונדריה מלאט מתחמצן לאוקסלואצטט על ידי האנזים מלאט דהידרוגנאז – malate dehydrogenase, תוך כדי המרת מולקולת +NAD ל – +NADH+ H.
מולקולת האוקסלואצטט עוברת טרנסאמינציה על ידי האנזים אספרטט אמינוטרספראז - aspartate aminotransferase ונוצרת מולקולת אספרטט - Aspartate. בריאקציית הטרנסאמינציה מועברת קבוצת האמינו מגלוטמט - Glutamate ונוצרת מולקולת אלפא קטוגלוטראט.
מולקולת האספרטט יוצאת אל המרווח הבין ממברנלי על ידי טרנספורטר – Glutamate – aspartate transporter, אשר מכניס אל המיטוכונדריה גלוטמט בכיוון הנגדי.
מולקולת האספרטט עוברת תהליך דאמינציה על ידי האנזים אספרטט אמינוטרספראז - aspartate aminotransferase ונוצרת מולקולת אוקסלואצטט אשר מתחילה את המחזור מחדש.
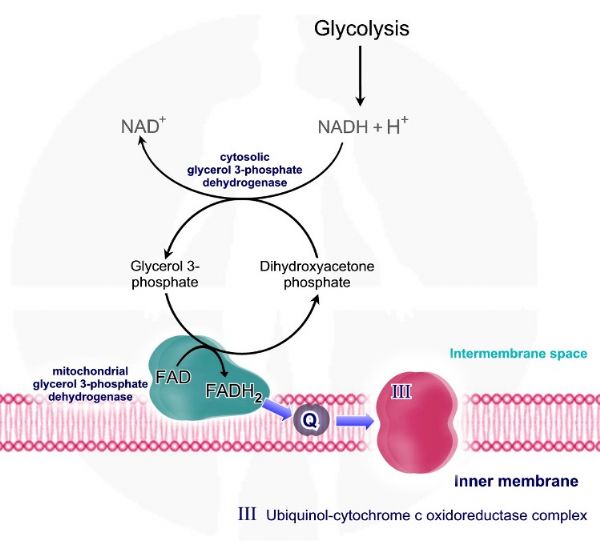
מעבורת הגליצרול -3 פוספט - Glycerol - 3 phosphate shuttle. דהידרוקסי אצטון פוספט - Dihydroxyacetone phosphate אשר נוצר בגליקוליזה מומר בציטוזול על ידי האנזים גליצרול - 3 פוספט דהידרוגנאז - Glycerol-3 phosphate dehydrogenase לגליצרול -3 פוספט - Glycerol - 3 phosphate. גליצרול - 3 פוספט יכול להיכנס אל המיטוכונדריה.
בממברנה הפנימית גליצרול – 3 פוספט מומר בחזרה לדהידרוקסי אצטון פוספט על ידי האנזים גליצרול - 3 פוספט דהידרוגנאז מיטוכונדריאלי או בשמו השני - פלאבו פרוטאין דהידרוגנאז flavoprotein dehydrogenase. אנזים זה תלוי ב – FAD כקו פקטור לשם העברת אלקטרונים. מתקבלת מולקולת FADH2 אשר ממשיכה את תהליך הובלת האלקטרונים. בדומה לקומפלקס -– II Succinate-Q oxidoreductase יכול לקשור אלקטרונים ישירות ל - ubiquinone – Q ללא הוצאת פרוטוני אל המרווח הבין ממברנלי.
מולקולת הדהידרוקסי אצטון פוספט חוזרת אל הציטוזול ומתחילה את המחזור מחדש.
F0 - החלק הטרנסממברנלי הבנוי מתעלה אשר דרכה חוזרים הפרוטונים - יוני +H אל המרווח התוך ממברנלי.
F1 - קומפלקס הבנוי מ - 6 תת יחידות, מסנתז ATP תוך כדי ניצול מפל הפרוטונים, ממיר אנרגיה כימית לאנרגיה מכאנית.